
相关文章链接:
1. 什么是单细胞测序
在前面我们介绍了测序技术的发展历史,以及测序技术的基本原理。在这一节,我们将真正的进入到单细胞测序的领域,介绍单细胞测序的基本原理,以及单细胞测序的应用。
在细胞的整个周期内,基因组、表观组、转录组和蛋白组是相互作用和相互调控的1。基因组和表观组的改变影响了转录本和蛋白的改变,而这些改变会反映到细胞水平的功能变化,同时又会反过来导致基因和表观遗传上的变化。通过理解整个复杂的调控机制,可以帮助我们更好的理解细胞的发育、分化、疾病和药物的作用等等。

让我们来想一下测序技术在这其中都可以做些什么:
- 基因组学相关,比如DNA的序列,单核苷酸多态性(Single Nucleotide Polymorphism,SNP),染色体拷贝数变异(Copy Number Variation,CNV),染色体结构变异(Structural Variation,SV),染色体构象分析(Hi-C,3C,4C,5C)等等
- 表观基因组学相关,比如DNA甲基化(DNA Methylation,包括5mC,5hmC,5fC,5caC等),DNA损伤和DNA修饰,组蛋白修饰,染色质可及性分析等
- RNA相关,比如RNA序列分析,RNA表达量分析,RNA二级结构分析,RNA splicing分析等等
- RNA表观遗传学相关,比如RNA editing,RNA修饰(经典的tRNA和rRNA修饰等),mRNA修饰(比如m6A,m1A, m5C,pesudouridine等)
- 蛋白组学检测,比如Olink公司的基于PEA技术的多重蛋白检测平台
- 其它各种应用2
而单细胞测序,不那么严谨的简单来说,就是在单个细胞的水平上去做上面各种应用。那么我们为什么要去在单细胞水平去做这些研究呢?一个核心点便是细胞之间的异质性(heterogeneity)。
比如从最初的一个受精卵开始,在发育的过程中,一生二,二生四,四生万物,最终分化出了不同的体细胞,细胞之间从外部形态到内部基因表达和蛋白表达差异巨大。又比如肿瘤的异质性,在肿瘤的生长过程中,经过多次的分裂增殖,肿瘤内部不同的细胞之间在基因或者转录组等分子生物学水平层面发生改变,导致这些细胞在生长速度、转移能力和抗药性等方面产生了差异。
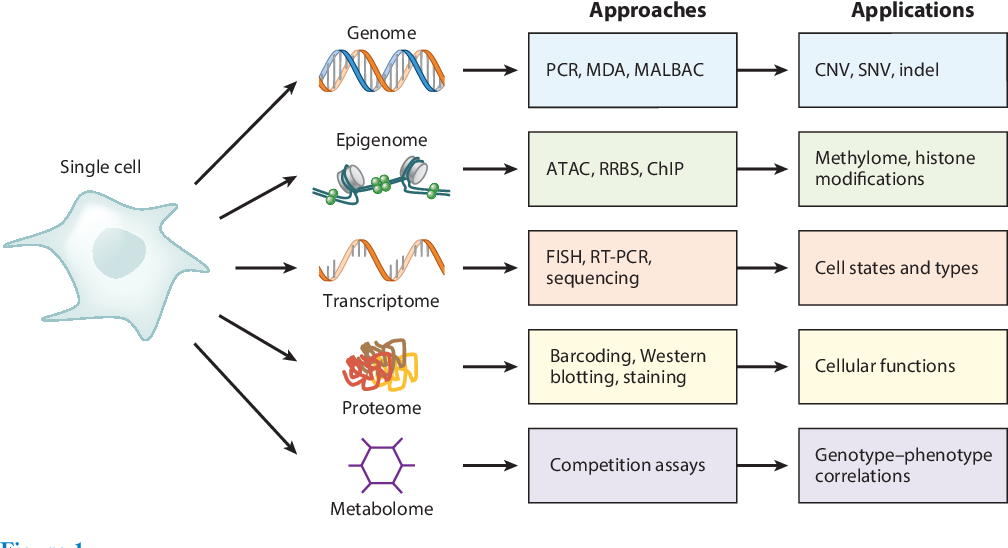
Deng, Y., et al. (2019). Single-Cell Omics Analyses Enabled by Microchip Technologies. Annual Review of Biomedical Engineering.
因此,传统的测序方法虽然提供了大量的基因组学或者转录组学的数据,但其最终的数据来自于大量细胞的信号的平均,通常反映的是单个组织或者单个个体的一个平均水平。而单细胞测序则可以提供来自于单个细胞的信号,这些信号可以反映出细胞之间的差异性,从而可以帮助我们更好的理解细胞之间的差异,更好的去探究疾病的发生和发展机制。
2013年,“单细胞测序技术”被Nature评为年度技术,2019年“单细胞多组学技术”被评为年度技术,其重要性可见一斑。单细胞技术的一个标志性应用是构建基因图谱,比如人类细胞图谱计划(Human Cell Atlas,HCA)等。
单细胞测序技术是一个比较大的概念,从测序的对象来看,可以分成单细胞基因组测序,单细胞转录组测序,单细胞蛋白组测序和单细胞多组学测序等,而近年来兴起的空间转录组学等也是单细胞测序的一个新方向。在这篇文章里,我们将主要讨论单细胞基因组测序的技术。
2. 单细胞基因组测序技术
单个细胞的DNA含量很少,只有5-10 pg左右,远低于常规的全基因组测序方法的最低样本投入量要求,因此,单细胞基因组测序技术需要解决由于DNA含量低而带来的以下几个难题:
- 如何实现DNA不同区域的均匀扩增,避免极少数区域的DNA扩增过多,造成少数区域的测序深度过高,而其他区域的测序深度过低的情况。
- 如何提高全基因组的覆盖率,避免由于常规建库过程中的DNA丢失导致基因组很多区域没有被测到。
- 如何提高建库扩增中的扩增效率,由于单细胞测序的DNA起始量很低,为了满足上机测序的要求,就需要进行非常高效率的DNA扩增,而这一点其实适合第一条相矛盾的,因为更多的扩增就意味着更多的扩增bias的积累。
为了解决这些方法,研究者们开发了多种全基因组扩增技术(Whole Genome Amplification,WGA),其发展历史可以参见图33。WGA技术按照技术路线的不同可以分为基于PCR的方法(如DOP-PCR),基于等温扩增的方法(如MDA),结合PCR和等温扩增的方法(如MALBAC)和基于转座酶的方法(如LIANTI,META-CS)等。而在这些技术中最为典型的就是DOP-PCR技术,MDA技术,MALBAC技术和LIANTI技术,下面我们将分别介绍一下这几个技术的技术原理。

2.1 Phi29 DNA Polymerase
在介绍MDA技术之前,让我们先来认识一下Phi29 DNA Polymerase(Φ29聚合酶)。Φ29聚合酶是从枯草芽孢杆菌Φ29噬菌体基因组中发现的4,并在1984年由Margarita Salas和Blanco教授进行了克隆表达5。
Φ29聚合酶的相对分子量为66kD,N端是核酸外切酶结构域,Asp-12、Glu-14、Asp-66或Asp-169的突变显示其核酸外切酶活性降低了100倍并且失去了链置换的活性;C端和其它DNA聚合酶类似,palm、fingers和thumb一起组成了聚合酶活性结构域。Φ29聚合酶是一个常温扩增酶,在30°C进行扩增反应,同时具有链置换的活性。
Φ29聚合酶的最广泛的应用是滚环扩增(Rolling-circle amplification,RCA)和多重链置换扩增(Multiple Displacement Amplification,MDA),常见图4。
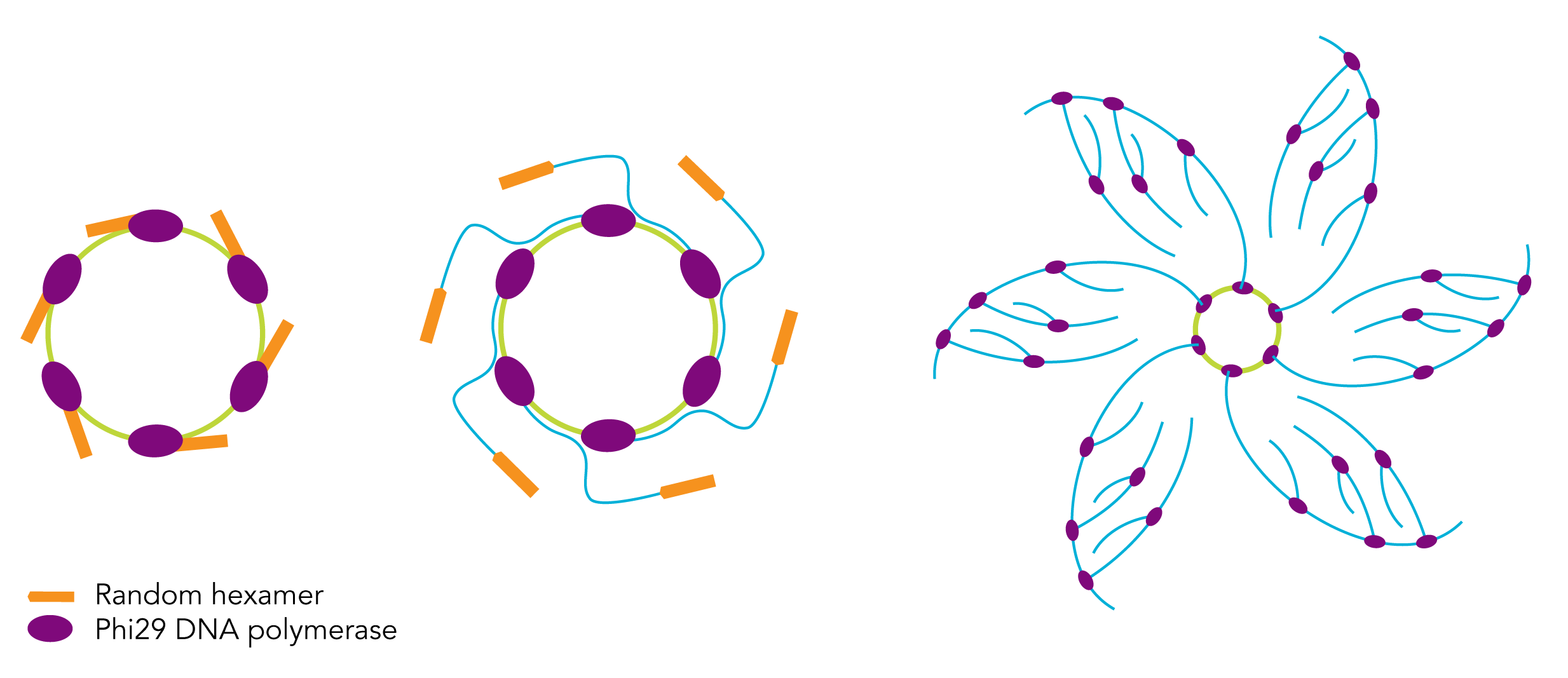
RCA是一种体外等温核酸扩增技术,其原理是Φ29聚合酶以环状DNA为模板,通过短的DNA引物(和部分环状模板互补),在酶的催化作用下合成DNA子链,同时由于Φ29聚合酶的链置换活性,在复制一圈后Φ29会把之前合成的互补链置换出去,最后形成了包含几十上百个重复的模板互补片段的序列。多重链置换则指的是在合成的过程中,新生链的合成和置换同时在多个地方同时进行,一条新生链的合成还未完全完成,这条链上又开始了新一轮的延伸和置换。
2.2 DOP-PCR技术
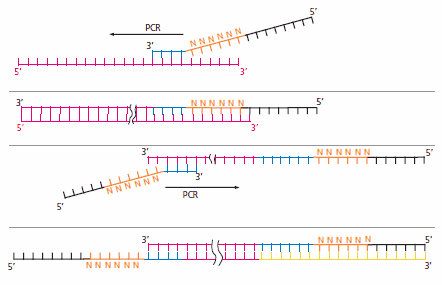
DOP-PCR技术(Degenerate Oligonucleotide Primed PCR)技术使用简并碱基引物,引物可以结合到DNA模板的任意部位,从而实现随机扩增得到全基因组序列。引物的5'端是正常的引物序列,3'端是6 nt的简并碱基(N~6~),3'端和DNA链随机结合并进行延伸6。
该方法的缺点是扩增后的基因组覆盖度不高,而这主要由其指数式扩增导致的扩增均一性差导致。
2.3 MDA技术
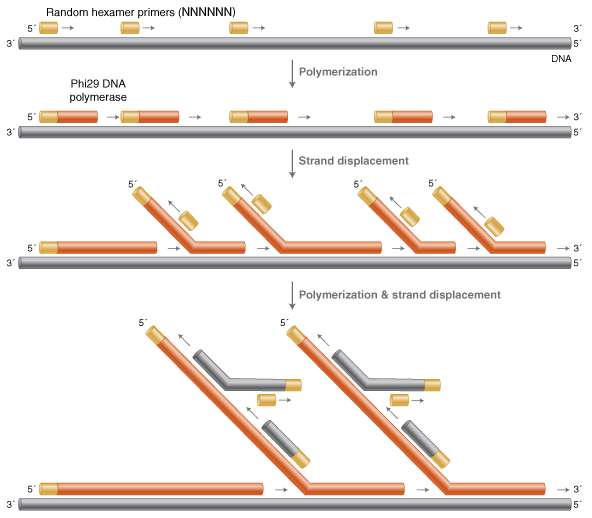
2001年,Laskin等发明了MDA技术,利用了刚才提到的Φ29聚合酶来进行,使用一条六个简并碱基的随机引物(N~6~)在恒温条件下与基因组随机退火,并在Φ29聚合酶的作用下发生多重链置换扩增反应,而置换产生的单链序列又可以和随机引物进行新一轮的延伸置换,从而最终可以合成长达50-100 kb的产物。同时由于Φ29聚合酶的3'-5'核酸外切酶活性和矫正活性,在这个扩增过程中还可以保证比较高的复制保真性。
MDA方法具有比较高的扩增效率和高保真性,在SNV的分析和大片段文库构建上有着显著的优势,而且实验操作简单,对实验条件要求低。MDA适用于各种样本类型,包括单个淋巴细胞、单个卵裂球等,还可以直接从生物样本里进行扩增,但是对于石蜡包埋固定的标本或者降解严重的DNA的扩增效果不佳。
但由于MDA法是一种直接指数扩增,因此存在着序列偏好性,而且这种偏好性不能重复,因此不适合进行CNV的分析。
为了克服扩增的bias缺陷,在MDA的基础上出现了ddMDA、eMDA和TruePrime等改良版技术。其中ddMDA和eMDA都是通过把扩增过程分散到数百万的小液滴中完成,从而达到提高扩增均一性和纠正bias的目的。TruePrime技术这是将MDA技术中的N~6~引物替换成一个以DNA为模板的引物合成酶来提高扩增均一性。
2.4 MALBAC技术
2012年,现任北京大学理学部主任、美国国家科学院院士谢晓亮教授团队在Science上连续发表了两篇应用其发明的新WGA技术进行单细胞测序的文章7,公布了一种名为“多次退火环状循环扩增技术”(Multiple Annealing and Looping-Based Amplification Cycles,MALBAC)的方法,并成功地实现了但细胞水平的SNV和CNV的研究8。
MALBAC结合了PCR和MDA两种技术,同时兼顾了基因组扩增过程中的保真性和均一性,并实现了基因组的高覆盖率。
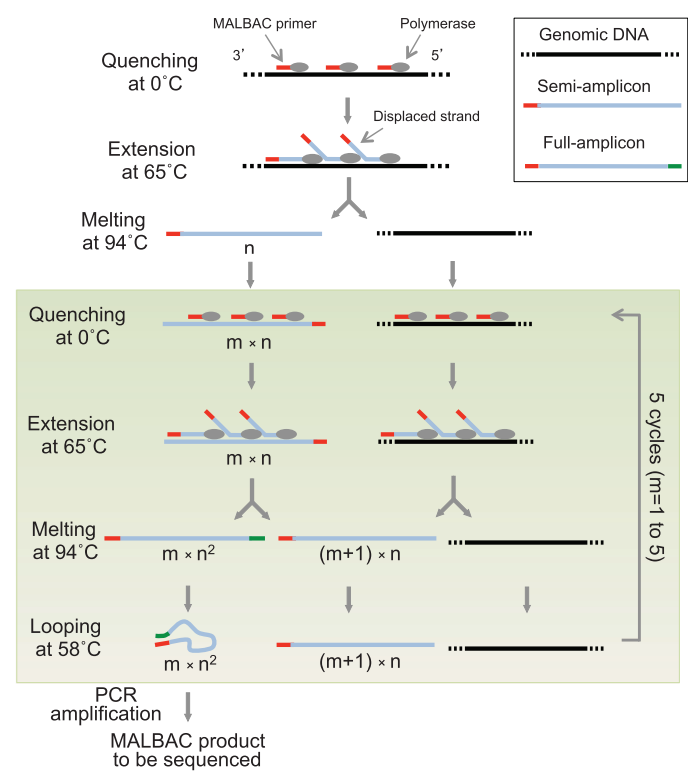
MALBAC的技术原理参见图7,其使用的35nt长的引物由一段固定的27 nt通用引物序列(GTG AGT GAT GGT TGA GGT AGT GTG GAG)和8 nt的随机简并碱基序列(N~8~)组成,在0°C的时候N~8~序列可以和模板DNA任意退火,升温到65°C之后使用同样具有链置换活性的BstDNA聚合酶进行延伸,得到一系列的半扩增子(Semi-amplicon),然后94°C解链得到单链DNA。
然后进入接下来的5轮线性扩增反应,在每一轮线性扩增中,首先在0°C退火和65度延伸,这是上一轮的半扩增子就形成了两端具有互补接口的全扩增子(Full-amplificon),随后在94°C解链后降低到58°C使得全扩增子的两端进MA行结合形成loop结构,从而避免了引物和末端结合导致全扩增子的自我指数扩增,从而保证该循环发生的是线性扩增。经过5轮线性扩增循环后,就可以得到大量的全扩增子作为接下来的PCR反应的模板,并使用27 nt的通用引物进行指数PCR反应。这时只有全扩增子可以得到有效的扩增。
在图6中,n代表了一个循环中,有多少个扩增引物可以配对到一个模板上,我们可以看到,全扩增子的产量是$mn^2$,也就是说,全扩增子的数量和循环数m成正比,而不是和$2^m$成正比,这样就可以保证全扩增子的产量是线性的,而不是指数的。同时,由于全扩增子的产量是线性的,所以MALBAC的扩增产量也是线性的,这样就可以保证MALBAC的均一性。
具有比MDA方法更为均衡的扩增结果,但是由于Bst聚合酶的保真度不如Φ29聚合酶,在进行单细胞基因型分析时,其假阳性率偏高。MALBAC技术在胚胎移植等领域的应用较多。
2.5 LIANTI技术
MDA技术和MALBAC技术均需要进行一步随机的PCR指数扩增,而只要进行指数扩增就会导致不均匀扩增,但为了得到足够多的DNA进行建库,我们好像又不得不进行指数扩增。
2017年,同样是谢晓亮教授,在Science上发表了其Linear Amplification via Transposon Insertion(LIANTI)技术9,这项技术在整个基因组的扩增过程中都是线性扩增技术。
首先,使用Tn5转座酶结合带有T7 promoter的LIANTI序列,然后使用该复合体,通过Tn5随机插入单细胞基因组DNA,再使用T7 RNA聚合酶进行体外转录获得线性扩增的RNA转录本,之后在通过逆转录和二链合成得到线性扩增的双链DNA模板,这样就得到了足够进行正常建库的DNA样本。在这个过程中,全程没有使用指数扩增的方法,保证了基因组的线性扩增。
LIANTI对于检测CNV的精确度非常的高,而且检测SNV的灵敏度相对于其它方法也大大提高,并且对于全基因组的覆盖率可以达到97%,ADO(Allele Dropout Rate)的比例也只有18%左右,低于MDA方法的30-40%和MALBC方法的45%。同时,由于使用了更高效的DNA聚合酶,LIANTI技术的单碱基扩增错误率被降到了$10^{-7}$一下。

3. 单细胞基因组测序技术的实际应用
WGA技术在医学上的实际应用,多在于辅助生殖领域。2014年9月19日,世界上首例经MALBAC测序进行单基因遗传病筛查的试管婴儿在北京大学第三医院诞生10,成为胚胎植入前遗传诊断技术的成功实施案例代表。
在这个案例中,男方为单基因显性遗传病患者,其后代无论男女均有1/2的概率患病,研究人员通过辅助生殖技术获得了18枚囊胚,然后利用显微操作技术得到了极少量的细胞,采用MALBAC技术进行检测,从中挑选出了3枚既不包含致病位点又不包含新发现的突变位点,同时染色体正常的胚胎,选择其中一枚移植到女方子宫内,最终经过羊水细胞基因验证和脐血基因检测均证实不含有致病位点。
目前世界上已经发现的单基因遗传疾病有7000多种,其中4000多种已经找到明确的致病基因。虽然大多数单基因遗传疾病在总人口中的发病率很低,但是因为种类极多,总患病人口数量相当庞大,给患者家庭和国家带来沉重的经济负担和生活负担。
胚胎遗传学诊断是一种早期的产前诊断技术,可有效避免中期被迫引产,有效地防止遗传病患儿的出生,同时彻底阻断遗传病在家族中的延续,对预防单基因遗传病的发生和传递具有非常重大的科学及社会意义。
相信随着WGA等技术的发展,未来将会有更多的临床应用成功案例的出现,切实地实现技术造福人类的目标。而这,也是我们做技术的初衷。

参考文献
-
Ogbeide, S., Giannese, F., Mincarelli, L., & Macaulay, I. C. (2022). Into the multiverse: advances in single-cell multiomic profiling. Trends in genetics : TIG, 38(8), 831–843. https://doi.org/10.1016/j.tig.2022.03.015 ↩
-
Khan, T., Becker, T. M., Po, J. W., Chua, W., & Ma, Y. (2022). Single-Circulating Tumor Cell Whole Genome Amplification to Unravel Cancer Heterogeneity and Actionable Biomarkers. International journal of molecular sciences, 23(15), 8386. https://doi.org/10.3390/ijms23158386 ↩
-
Salas M. (2007). 40 years with bacteriophage ø29. Annual review of microbiology, 61, 1–22. https://doi.org/10.1146/annurev.micro.61.080706.093415 ↩
-
Telenius, H., Carter, N. P., Bebb, C. E., Nordenskjöld, M., Ponder, B. A., & Tunnacliffe, A. (1992). Degenerate oligonucleotide-primed PCR: general amplification of target DNA by a single degenerate primer. Genomics, 13(3), 718–725. https://doi.org/10.1016/0888-7543(92)90147-k ↩
-
Zong, C., Lu, S., Chapman, A. R., & Xie, X. S. (2012). Genome-wide detection of single-nucleotide and copy-number variations of a single human cell. Science (New York, N.Y.), 338(6114), 1622–1626. https://doi.org/10.1126/science.1229164 ↩
-
Lu, S., Zong, C., Fan, W., Yang, M., Li, J., Chapman, A. R., Zhu, P., Hu, X., Xu, L., Yan, L., Bai, F., Qiao, J., Tang, F., Li, R., & Xie, X. S. (2012). Probing meiotic recombination and aneuploidy of single sperm cells by whole-genome sequencing. Science (New York, N.Y.), 338(6114), 1627–1630. https://doi.org/10.1126/science.1229112 ↩
-
Chen, C., Xing, D., Tan, L., Li, H., Zhou, G., Huang, L., & Xie, X. S. (2017). Single-cell whole-genome analyses by Linear Amplification via Transposon Insertion (LIANTI). Science (New York, N.Y.), 356(6334), 189–194. https://doi.org/10.1126/science.aak9787 ↩
Comments NOTHING